Modernizing gut-brain axis research in nutritional Science: The role of human-centered New Approach Methodologies
Abstract
Background
The gut-brain axis is a complex communication network that connects the gastrointestinal system with the central nervous system, significantly influencing various health outcomes, such as mental health, cognitive function, metabolic regulation, and immune responses. While traditional research models, particularly animal studies, have provided valuable insights, they often overlook the intricate and human-specific interactions within this axis. Consequently, translating findings from these models into clinical applications has been challenging. However, recent advancements in human-based Novel Approach Methodologies (NAMs), like organoids, organs-on-chip, and omic sciences, present innovative tools for investigating the gut-brain axis with improved accuracy and relevance to human physiology. These methodologies facilitate a deeper understanding of the molecular and cellular mechanisms by which nutritional interventions affect not only mental health but also a wider range of gut-brain-related health outcomes. Scope and approach: Scope and approach: This paper explores how NAMs are revolutionizing gut-brain axis research by providing more accurate models that replicate human physiology, thereby replacing less effective traditional approaches.
Key findings and conclusion
By using these advanced methods, researchers can produce detailed data that better mirror human responses to dietary components, resulting in more effective and personalized strategies for managing and enhancing gut-brain health. Future research should concentrate on utilizing NAMs to deepen our understanding of the gut-brain axis in nutritional science, which will ultimately lead to more targeted and effective health interventions for various conditions.
Previous article in issue
Next article in issue
Keywords
Gut-brain axis
Nutrition
Organoids
Organs-on-chip
Omics
1. Introduction
The gut-brain axis is a complex and dynamic communication network that intricately links the gastrointestinal system with the central nervous system, playing a crucial role in regulating various health outcomes, including mental health, cognitive function, metabolic regulation, and immune responses. The increasing recognition of this axis as a central player in mediating the effects of diet on health has spurred significant interest in understanding how nutritional interventions can influence these interactions (Ribeiro et al., 2022).
Historically, research into the gut-brain axis has relied heavily on simplistic in vitro systems or animal models (Moysidou & Owens, 2021). While these models have provided foundational insights into the basic mechanisms of gut-brain communication, they also present significant limitations. Simplistic in vitro models, which have been used to study isolated aspects of the gut-brain axis, are often too reductionist to capture the complex and multifaceted interactions that occur in a living organism. Although useful for certain types of mechanistic studies, these models frequently lack the necessary physiological relevance to accurately represent the intricacies of human biology. On the other hand, the differences in microbiota composition, immune system responses, and neural architecture between humans and commonly used animal models often limit the direct applicability of research findings to human health (Hartung, 2024). Moreover, the ethical concerns associated with animal research have become increasingly prominent, leading to a growing demand for alternative methods that reduce or replace the use of animals in scientific studies. The ethical imperative to reduce the use of animals in research is supported by the principles of the 3 R s: Replacement, Reduction, and Refinement. These principles advocate for the replacement of animal models with non-animal methods wherever possible, the reduction in the number of animals used, and the refinement of experimental techniques to minimize suffering. In response to these concerns and the limitations of traditional models, there has been a significant push towards developing more human-relevant research methodologies (Pound & Ritskes-Hoitinga, 2018). Advancements in New Approach Methodologies (NAMs), such as organoids, organs-on-chip, and omic sciences, have provided researchers with powerful tools to study the gut-brain axis in ways that are not only more applicable to human biology but also align with the ethical goals of reducing reliance on animal models (Moysidou & Owens, 2021; Reardon, 2024; Sheng et al., 2023; Zhang et al., 2024). The human gut microbiome has emerged as a key player in the bidirectional communication of the gut-brain axis, affecting various aspects of homeostasis and pathophysiology. Until recently, the majority of studies that seek to explore the mechanisms underlying the microbiome-gut-brain axis cross-talk, relied almost exclusively on animal models, and particularly gnotobiotic mice (Moysidou & Owens, 2021). Despite the great progress made with these models, various limitations, including ethical considerations and interspecies differences that limit the translatability of data to human systems, pushed researchers to seek for alternatives. Over the past decades, the field of in vitro modelling of tissues has experienced tremendous growth, thanks to advances in 3D cell biology, materials, science and bioengineering, pushing further the borders of our ability to more faithfully emulate the in vivo situation. The discovery of human induced pluripotent stem cells (iPSCs) has offered a new source of cells, while their use in generating gastrointestinal and brain organoids, among other tissues, has enabled the development of novel 3D tissues that better mimic the native tissue structure and function, compared with traditional assays (Hall & Bendtsen, 2023). In parallel, organs-on-chips technology and bioengineered tissues have emerged as highly promising alternatives to animal models for a wide range of applications (Ingber, 2020). Here, we discuss how recent advances and trends in this area can be applied in host-microbe and host-pathogen interaction studies. In addition, we highlight paradigm shifts in engineering more robust human microbiome-gut-brain axis models and their potential to expand our understanding of this complex system and hence explore novel, microbiome-based therapeutic approaches. These methodologies enable the exploration of the molecular and cellular mechanisms underlying gut-brain communication with greater precision and relevance, thereby facilitating the development of more targeted and effective nutritional interventions. This paper aims to explore the transformative potential of NAMs in modernizing gut-brain axis research, particularly within the context of nutritional science.
2. The gut-brain axis: mechanisms and health implications
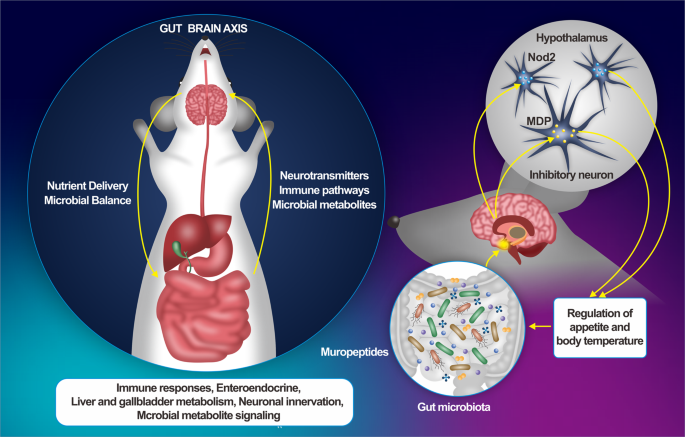
The gut-brain axis (GBA) is a multifaceted communication network that connects the gastrointestinal system with the central nervous system (CNS), playing a critical role in regulating various aspects of health. This axis functions through several key pathways, including neural, endocrine, immune, and metabolic routes, and is significantly influenced by the gut microbiota (Fig. 1) (Macpherson et al., 2023). The brain’s vasculature forms the blood-brain barrier (BBB), which is more restrictive than the systemic vasculature due to the neurovascular unit (NVU). This unit consists of microvascular endothelial cells, pericytes, microglia, and astrocytes. The BBB protects brain tissues from harmful substances while allowing essential molecules like neurotransmitters, oxygen, and glucose to pass through. Transport across this selectively permeable barrier is facilitated by both active and passive mechanisms, which are regulated by the NVU’s specialized cells, surrounding tissues, and blood flow-induced shear stress. The integrity of this barrier is crucial for the efficient passage of molecules (McConnell & Mishra, 2022). The intestinal epithelium protects the body’s circulatory system from harmful foreign substances. Compromise of the gut barrier can result in a condition referred to as “leaky gut,” in which harmful substances, such as toxins, pathogens, and inflammatory molecules, may translocate into the bloodstream. This breach of the gut barrier can trigger systemic inflammation, which has the potential to affect the central nervous system, thereby influencing brain function and contributing to the development of neurological and psychiatric disorders (Vancamelbeke & Vermeire, 2017).
Download: Download high-res image (840KB)
Download: Download full-size image
Fig. 1. Overview of the gut-brain axis (GBA): The GBA involves complex connections between the gut and brain, separated by the gut barrier and the blood-brain barrier (BBB). The gut microbiome plays a critical role by producing molecules such as immune signals, metabolites, and neurotransmitters. The enteric nervous system, known as the “second brain,” transmits signals to the central nervous system (CNS), alongside vagal and endocrine pathways. The BBB, composed of endothelial cells, pericytes, and astrocytes, further highlights the GBA’s physiological complexity, important in both health and disease. Abbreviations: Astr, Astrocyte; B, Lymphocyte B; BV, Blood Vessel; CNS, Central Nervous System; DC, Dendritic Cell; Dopa, Dopamine; EEc, Enteroendocrine Cell; Epi, Epinephrine; EpC, Epithelial Cell; Ec, Endothelial Cell; GABA, Gamma-Amino-Butyric Acid; Gc, Goblet Cell; GM, Gut Microbiota; Ig, Immunoglobulin; Mac, Macrophage; Mc, Microfold Cell; NE, Norepinephrine; Pan, Paneth Cell; Per, Pericyte; SCFAs, Short-Chain Fatty Acids; Tg, Tight Junction; T, Lymphocyte T; 5HT, 5-Hydroxytryptamine (Serotonin). Adapted from (Hall & Bendtsen, 2023) Creative Commons Attribution License (CC BY).
While the brain and gut are distinct compartments, each separated by their own barriers – the gut barrier and the BBB – they are connected by a bidirectional communication network that facilitates signals between the gut and the brain. This concept aligns with a holobiontic view, where the body operates as a host in a symbiotic relationship with the gut microbiome, contributing to immunity, nutrition, and overall health (Reynoso-García et al., 2022). A growing body of epidemiological and pre-clinical studies suggests that an unhealthy gut is linked to various brain disorders and mental health conditions, though the precise mechanisms remain unclear. The GBA has been associated with several neurodevelopmental, functional, and psychiatric conditions, including multiple sclerosis, neurodegenerative diseases like Alzheimer’s disease and Parkinson’s disease, as well as major depressive and mood disorders (Sorboni et al., 2022). Epidemiological research further highlights that individuals suffering from these brain-related disorders often have a dysbiotic gut microbiome, characterized by reduced microbial richness (alpha diversity) and altered microbial composition (beta diversity) when compared to healthy individuals. A recent meta-analysis involving psychiatric patients and control participants demonstrated a consistent pattern across individuals with major depressive disorder, bipolar disorder, psychosis, schizophrenia, and anxiety. The study found that specific anti-inflammatory, butyrate-producing gut bacteria were depleted, while pro-inflammatory bacteria showed an increase (Nikolova et al., 2021).
The production of neurotransmitters and neuroactive compounds, such as short-chain fatty acids (SCFAs), by the gut microbiota is essential for maintaining CNS homeostasis (Ashique et al., 2024).
Dietary components and biologically active compounds derived from food have the potential to influence gut microbiota composition and abundance (Rinninella et al., 2023), gut barrier function, and systemic inflammation, thereby affecting brain health and overall well-being (Cassotta et al., 2020; Hao et al., 2020; He et al., 2020; Li, Huang, et al., 2023; Wang et al., 2022). Understanding the mechanisms underlying the complex communication within the gut-brain axis provides a foundation for developing targeted nutritional interventions aimed at improving both mental and physical health (Ashique et al., 2024). As research in this area continues to grow, it is becoming increasingly evident that the GBA plays a central role in many aspects of health, making it a key focus for future nutritional research and therapeutic development.
3. Limitations of traditional gut-brain axis models
Traditional models used to study the gut-brain axis – primarily animal models and basic in vitro systems – face significant limitations that hinder their ability to accurately replicate human physiology and predict clinical outcomes (Schellekens et al., 2023). The main limitations of traditional in vivo (animal) and in vitro models in GBA research are summarized in Table 1.
Table 1. An overview of the main limitations of traditional in vivo and in vitro models in GBA research.
Limitations of Animal Models
Limitations of Traditional In Vitro Models
Differences in microbiota composition, abundance, and functional capacities (Nagpal et al., 2018)
Lack of mechanical signals from a 3D environment, flow, and movement (Hartung, 2007)
Divergences in the immune system (Khanna & Burrows, 2011)
Lack of biochemical cross-talk with other cell types (Hartung, 2007)
Anatomical and physiological differences in the gastrointestinal tract
Lack of the three-dimensional architecture and diverse cell types found in the gut or brain environment
Behavioral and cognitive differences compared to humans, differences in neuroanatomy and brain function (Jeong et al., 2021; MacLean, 2016)
Static environment (Hartung, 2007)
Different nutritional physiology and behavior (Bogatyrev et al., 2020; Gregor et al., 2020)
Absence of systemic responses
Variability in ADMET (absorption, distribution, metabolism, excretion, and toxicity) of food-derived bioactive compounds (Toutain et al., 2010)
Co-culturing mammalian and gut-microbiota bacterial cells poses significant challenges due to their different culture requirements (Davenport Huyer & Radisic, 2021).
Artificial conditions in laboratory environments and routine-induced stress (Bailey, 2017)
Non-human or tumor-derived cell origins (Estévez-Priego et al., 2023; Hartung, 2007)
Ethical concerns regarding animal suffering
Waste accumulation, limited oxygen and nutrient supply (Hartung, 2007)
Low translational relevance to human contexts (Hartung, 2024; Schellekens et al., 2023)
Cellular stress and altered behavior (Hartung, 2007)
Requiring significant resources: time, money, and labour
Limited relevance to real human conditions
3.1. Animal models and their limitations
Due to its complexity, research on the GBA has traditionally relied on in vivo animal models (Raimondi et al., 2019). Various animal species have been utilized as models in research on the microbiota-gut-brain axis and nutritional sciences. Among mammals, rodents, particularly mice, have been extensively used to initially explore the microbiota’s role in physiological processes (Mayer et al., 2022). For instance, until recently, most studies investigating the mechanisms behind the microbiome–gut–brain axis relied almost exclusively on gnotobiotic mice. Other animals, such as, honeybees (Chang et al., 2022), zebrafish (Chen et al., 2021), pigs (Sutkus et al., 2022), Caenorhabditis elegans (D. H. Kim & Flavell, 2020), dogs (Kubinyi et al., 2020), and rhesus macaques (Johnson et al., 2022), have also been employed in microbiota-gut-brain axis studies.
Although animal models have provided valuable insights into basic biological mechanisms, several inherent differences between species limit the applicability of these findings to humans (Han & Jang, 2022).
•
Microbiota Composition: The gut microbiota composition in animals differs significantly from that in humans. These differences extend not only to the types of microbial species present but also to their functional capacities. For example, certain microbial strains that are common in humans may be absent or rare in rodents, leading to different metabolic outputs that can affect gut-brain signaling in ways that do not translate to human physiology (Bogatyrev et al., 2020; Nagpal et al., 2018). Moreover, the nutritional composition of commercially available rodent diets influences gut microbiota profiles, significantly affecting the reproducibility of results across different laboratories (Tuck et al., 2020).
•
Immune System Function: The immune system is another area where significant differences exist between species. Rodents have a different immune response profile compared to humans, particularly in the gut-associated lymphoid tissue (GALT). These differences can lead to variations in how the immune system interacts with the gut microbiota and responds to inflammation, affecting the interpretation of data on gut-brain interactions (Khanna & Burrows, 2011; Mestas & Hughes, 2004; St Clair, 2008).
•
Although the central nervous system’s basic structure is conserved across mammals, significant differences in neuroanatomy and brain function exist between animal models and humans, including non-human primates. These distinctions affect how the brain interprets signals from the gut, particularly in regions associated with higher cognitive functions and emotional regulation, which are more complex and highly developed in humans (Jeong et al., 2021).
•
Behavioral and Cognitive Differences: The behavioral repertoire of animal models is much simpler compared to that of humans, making it challenging to study complex psychological phenomena such as anxiety, depression, and cognitive decline. Additionally, the responses of animals to stress and other environmental factors can differ significantly from those of humans, limiting the relevance of findings to human conditions (MacLean, 2016).
•
Nutritional physiology and behaviour: There are considerable differences between animal models and humans regarding nutrition, dietary behavior, and the physiological effects of food-derived compounds. Species-specific variations in gastrointestinal physiology and metabolic rates can lead to distinct differences in nutrient absorption, metabolism, and overall bioavailability of dietary components. Additionally, the feeding behaviors and dietary preferences of animals often do not accurately reflect human eating patterns, which can further complicate the translation of findings from animal studies to human contexts (Bogatyrev et al., 2020; Gregor et al., 2020). Furthermore, the pharmacokinetics and pharmacodynamics of food bioactive compounds may differ significantly between species due to variations in metabolic pathways and receptor distribution (Toutain et al., 2010).
•
Laboratory conditions differ markedly from animals’ natural habitats, potentially limiting the generalizability of experimental findings. Animals in controlled lab environments are exposed to unique lifestyle factors, including a standardized diet, restricted physical and social interactions, and potential stressors from laboratory routines. These factors can influence both physiology and behaviour, making it challenging to fully translate laboratory results even to the same species in the wild. Consequently, these differences complicate the extrapolation of experimental data to real-world applications (Bailey, 2017).
The ethical and practical limitations of animal models also warrant consideration.
•
The use of animal models raises significant ethical issues, particularly regarding animal welfare.
•
The principles of the 3 R s (Replacement, Reduction, and Refinement) advocate for the reduction of animal use in research and the replacement of animal models with alternative methods wherever possible. Traditional models often fail to align with these ethical guidelines, especially when non-animal alternatives could provide more relevant data.
•
Animal studies are resource-intensive, requiring significant time, money, and labour. This can limit the scale and scope of research projects, especially in exploratory studies where large datasets are needed to uncover novel interactions within the gut-brain axis.
3.2. Inadequacies of basic in vitro models
Basic in vitro models, such as monolayer cultures of intestinal epithelial cells or isolated neuronal cells, are commonly used to study specific aspects of the gut-brain axis. While these models offer controlled environments for mechanistic studies, they have several limitations.
An important limitation is the simplistic nature of traditional cell cultures. Monolayer cultures do not capture the full complexity of the gut or brain environment. They lack the three-dimensional architecture and diverse cell types found in these organs, which are crucial for maintaining physiological functions. For example, the interactions between different cell types, such as enterocytes, immune cells, and neurons, are essential for accurate modeling of the gut-brain axis.
Co-culturing mammalian and gut-microbiota bacterial cells poses significant challenges due to their fundamentally different culture requirements. Aerobic bacteria, for example, tend to proliferate rapidly and can easily outcompete mammalian cells when grown on shared nutrient sources like glucose. On the other hand, anaerobic bacteria, such as those found in the human gut microbiome, are highly sensitive to oxygen and cannot be cultured under the standard conditions that are suitable for mammalian cells. This divergence in environmental needs makes maintaining a stable co-culture of both cell types particularly difficult (Davenport Huyer & Radisic, 2021). As a consequence, traditional in vitro models often exclude the gut microbiota, which plays a critical role in gut-brain communication. Without the microbiota, these models cannot replicate the production of microbial metabolites, such as short-chain fatty acids (SCFAs) and neurotransmitters, which are key mediators of gut-brain interactions.
Another significant drawback is that these systems are often based on cell lines derived from tumors or non-human sources. Tumor-derived cell lines, while convenient and easy to maintain, do not always exhibit normal cellular behaviour and may have altered metabolic and signaling pathways that do not accurately represent healthy tissue (Hartung, 2007). Similarly, the use of non-human cells can limit the translatability of findings to human physiology, as there may be species-specific differences in cellular responses and interactions (Estévez-Priego et al., 2023). Moreover, basic in vitro models are typically short-lived, limiting their ability to study long-term interactions and chronic conditions. The lack of continuous nutrient supply and waste removal in these systems often leads to cellular stress and altered behaviour, which can skew results and reduce their relevance to in vivo conditions.
3.3. Implications for research and therapeutic development
The limitations of traditional gut-brain axis models have significant implications for both research and therapeutic development. Inaccurate modeling can lead to misleading conclusions about the mechanisms underlying gut-brain interactions and the effects of food-derived compounds, thereby hindering the development of effective prevention measures or treatments for conditions such as depression, anxiety, and neurodegenerative diseases. Furthermore, the inability of these models to replicate human physiology accurately contributes to the high attrition rates observed in clinical trials, where promising preclinical results often fail to translate into effective therapies.
Despite the extensive use of animals in non-clinical research, a surprisingly small percentage of this research successfully translates to human clinical trials, with approximately 90 % of drug candidates failing in Phase 1 (Seyhan, 2019). This translational gap underscores the need for more human-relevant models that can better predict how interventions will work in clinical settings.
4. NAMs in gut-brain axis research
NAMs refer to innovative, non-animal testing methods and technologies that have emerged as powerful tools in biomedical research. These methodologies encompass a range of advanced techniques, including organoids, organs-on-chip, computational and omic sciences, which allow for the study of biological processes with a level of precision and relevance that is difficult to achieve with traditional models, particularly animal studies (Interagency Modeling and Analysis Group (IMAG), 2024). NAMs are designed to more closely model human biology, offering valuable insights into physiological and pathological processes. This section provides an overview of the key NAMs that are particularly suited for studying the GBA and investigating the impact of dietary bioactive compounds on gut-brain interactions.
4.1. Organoids: modeling human-specific gut and brain physiology
Organoids are three-dimensional, simplified, and miniature versions of organs grown in vitro from stem cells. These structures mimic key aspects of the architecture and function of their respective organs, although they are not fully developed or completely functional as the organs in vivo. Organoids are derived from pluripotent stem cells – either embryonic stem cells or induced pluripotent stem cells (iPSCs) – or adult stem cells, which self-organize into organ-like structures under appropriate culture conditions.
iPSCs can be generated from readily accessible somatic cells, such as skin-derived fibroblasts, blood-derived erythroblasts, or urine-derived epithelial cells (Raab et al., 2014), without the need for embryo destruction, thus avoiding associated ethical concerns. Furthermore, iPSCs generation does not rely on biopsy material obtained through invasive endoscopic procedures, facilitating the collection of source material from both healthy individuals and patients. This allows for the study and comparison of different genetic backgrounds. Patient-specific iPSCs offer an unlimited supply of disease-relevant cells tailored to the individual, providing access to previously inaccessible cell types, such as neurons, in a personalized manner. The ability of these cells to differentiate and organize into complex tissues with multiple cell types allows organoids to recapitulate many aspects of organ development, disease pathology, and physiology. Organoids, in particular gut or gastrointestinal (GI) and brain organoids are increasingly being used to study the gut-brain axis.
4.1.1. Gut organoids
GI or gut organoids replicate the cellular and structural features of the intestinal epithelium, including the presence of various cell types such as enterocytes, goblet cells, and enteroendocrine cells. By exposing gut organoids to different dietary components, researchers can study how specific nutrients affect intestinal cell function, gut-barrier function, gut microbiota composition, and the production of metabolites. The presence of various functional enteroendocrine cells in GI organoids, particularly enterochromaffin cells, which have been shown to activate the enteric nervous system (ENS) and transmit chemosensory signals to the brain, enables the modeling of aspects of the gut-brain axis communication (Hall & Bendtsen, 2023).
Pearce et al. (2020) demonstrated that microbiota-derived SCFAs, such as butyrate and propionate, modulate intestinal stem cell activity, promote epithelial differentiation, and enhance barrier function in human small intestinal organoid models (Pearce et al., 2020).
Danhof et al. (2023) used human intestinal organoids to show that the intestinal epithelium is capable of producing oxytocin. They found that Lactobacillus reuteri facilitates oxytocin secretion from both human intestinal tissue and organoids. Furthermore, their research demonstrated that this stimulation of oxytocin secretion is dependent on secretin, a gut hormone produced by enteroendocrine cells, while oxytocin is produced by enterocytes. Oxytocin, traditionally known for its roles in social bonding and stress regulation in the brain, is now being identified as an intestinal hormone that may influence gut-brain communication. The production of oxytocin by enterocytes in response to stimuli from gut microbes like Lactobacillus reuteri highlights the possibility that gut microbiota can modulate not only gut health but also brain-related functions through oxytocin-mediated pathways (Danhof et al., 2023).
Since the organoid lumen is enclosed in the construct centre and the basal membrane is exposed outwards, a common technique used to deliver microbiota or their metabolites to the apical surface of the epithelium is microinjection (Williamson et al., 2018). With this procedure, the internal environment need by microbes is maintained and is more appropriate for examining long-term relations with pathogens or commensals that typically infect the host from the lumen. However, it requires special equipment, and it is quite challenging to perform reproducible and quantitative experiments, while damage of organoids during the process is common. In an attempt to overcome these challenges, Co and collaborators (Co et al., 2019) developed a technique to reverse the polarity of enteroids, to expose their apical surface to the media without affecting the intestinal constructs structure and function, and efficaciously used this to recognise the infection patterns of enteropathogens. Otherwise, GI organoids are regularly grown before the enzymatic dissociation and then seeded onto Matrigel/ECM coated dishes or Transwell culture inserts, where they are able to create monolayers that comply with the epithelial barrier dynamics and allow for exposure of the apical/luminal surface to microbes or their metabolites, added in the culture medium (Hill & Spence, 2017). Even if these monolayers present the same cellular diversity of the organoids, they fail to capture the 3D microenvironment of the native tissue and may not be suitable for long-term experiments.
Even though GI organoids hold great promise as in vitro human models, their use in studies of microbiome-gut-brain signaling pathways is hindered by several limitations. These include difficulties in replicating age-related structural and functional features of native tissue, batch-to-batch size variability, and, most critically, the absence of essential components such as vasculature, the ENS, and the immune system. To overcome these challenges, researchers are focusing on developing co-cultures of GI organoids with immune cells and/or enteric neurons (Noel et al., 2017; Workman et al., 2017).
A 2017 study utilizing gut-mimicking organoids provided novel insights into the role of enterochromaffin cells (ECs) in detecting noxious substances, generating electrical impulses in nearby nerve fibers, and relaying critical information to the brain (Hampton, 2017). Through single-cell measurements within these organoids, the researchers identified key biophysical, pharmacological, and genetic properties of ECs. The data revealed that ECs are electrically excitable and can modulate serotonin-sensitive nerve fibers via synaptic connections. A comprehensive screening of gut-derived compounds, including dietary irritants such as allyl isothiocyanate, bacterial metabolites like isovalerate, and catecholamines (epinephrine, norepinephrine, dopamine), consistently activated ECs. Notably, these compounds have been implicated in the pathophysiology of irritable bowel syndrome. Furthermore, the study elucidated specific chemosensory receptors and signaling pathways that regulate EC activation and serotonin release in response to these stimuli. Ongoing research using intestinal organoids aims to further investigate how gastrointestinal disorders may alter the prevalence, function, and neural communication of ECs (Hampton, 2017).
Llorente (2024) recently emphasized the need for innovative co-culture models that integrate the ENS with intestinal organoids to improve gastrointestinal disease modeling and treatment approaches. The article discusses a companion protocol detailing methods for isolating and co-culturing myenteric and submucosal neurons with intestinal organoids. This model enables the study of ENS-intestinal interactions, offering a valuable tool not only for exploring motility disorders and other GI-related diseases but also for investigating the gut-brain axis (Llorente, 2024).
4.1.2. Brain organoids
Brain organoids have been utilized to investigate interactions between the neuro-immune and neuro-endocrine systems (Chukwurah et al., 2019), as well as host-microbe interactions involving pathogens such as Zika virus (Xu et al., 2019) and Toxoplasma gondii (Seo et al., 2020). Advancements in iPSC-derived GI, ENS, and brain organoids, along with vagal nerve neurons and enhanced co-culture techniques, could provide valuable models for studying the gut-brain axis and the role of microbiota in various diseases and health conditions (Chukwurah et al., 2019).
Human midbrain organoids have been successfully used to study the connection between aging and the gut microbiota-derived metabolite trimethylamine N-oxide (TMAO), which is produced from l-carnitine, choline, phosphatidylcholine, and betaine—compounds abundant in foods such as dairy products, egg yolks, and meat. Midbrain organoids treated with TMAO showed molecular changes related to aging, including elevated senescence marker expression and epigenetic modifications. Additionally, these TMAO-treated organoids exhibited neurodegenerative features, such as the loss of dopaminergic neurons, neuromelanin accumulation, and alterations in α-synuclein and Tau proteins. These findings suggest that TMAO plays a role in midbrain aging and pathogenesis, providing insights into how diet or intestinal dysfunction might increase the risk of neurodegenerative diseases like Parkinson’s (Lee et al., 2022).
A key limitation of organoids is their inability to replicate the physiological inter-organ communication required to reflect systemic responses. Despite this, the range of organoid applications is expanding rapidly, with significant progress being made toward developing more complex and sophisticated organoid-based model systems. For example, Trapecar et al. (2021) established a mesofluidic platform to explore gut-liver-cerebral interactions in the context of Parkinson’s disease (PD). The platform connected primary human gut and liver organoids with a cerebral organoid derived from human induced pluripotent stem cells within a microphysiological system, circulating a common culture medium containing human CD4+ regulatory T cells and T helper 17 cells. Using patient-derived cerebral organoids carrying the PD-causing A53T mutation, the study demonstrated that systemic interactions within the platform enhanced the in vivo-like behavior of the cerebral organoids and that microbiome-derived SCFAs increased the expression of pathways associated with PD pathology. This approach provided key insights into the systemic dynamics contributing to neurodegenerative processes in PD, highlighting the significant role of gut-liver-brain communication and the microbiome in disease progression (Trapecar et al., 2021).
Researchers have recently developed a model of the blood-brain barrier (BBB) using assembloids -combinations of organoids mimicking complex tissues. This model integrates brain and blood-vessel organoids, allowing the vessels to form capillary networks within the brain tissue, closely replicating the structure of the human BBB. The model successfully recreates both normal BBB function and genetic diseases associated with BBB defects, offering a promising tool for studying the BBB physiopathology (Dao et al., 2024). Organs-on-chip technologies provide a complementary solution by offering enhanced control over fluid flow, nutrient delivery, and mechanical forces, effectively addressing many of the limitations associated with organoids.
Some examples of applications of organoids in gut-brain axis research within the field of nutrition are summarized in Table 2.
Table 2. Some examples of organoid applications in GBA research in the field of nutrition.
Organoid Type
Examples of Applications
Bibliography
Gut Organoids
–
Replicating cellular and structural features of the intestinal epithelium, with diverse cell types such as enterocytes, goblet cells, and enteroendocrine cells.
–
Studying nutrient effects on intestinal cell functions, gut-barrier integrity, and microbial composition.
–
Investigating how specific dietary components influence microbiota composition and metabolite production.
(Hall & Bendtsen, 2023);
Study of microbiota-derived SCFAs effects on intestinal stem cell activity, epithelial differentiation, and barrier function
(Pearce et al., 2020)
Investigation of oxytocin production in response to gut microbiota and its influence on gut-brain communication
(Danhof et al., 2023)
Evaluation of gut-microbe interactions through microinjection or co-culture techniques
(Co et al., 2019; Williamson et al., 2018)
Research on ENS and immune cell co-cultures with GI organoids to model the gut-brain axis
(Noel et al., 2017; Workman et al., 2017) Hampton (2017)
Brain Organoids
Examination of neuro-immune and neuro-endocrine interactions, as well as host-microbe interactions with pathogens
(Chukwurah et al., 2019; Xu et al., 2019); (Seo et al., 2020);
Investigation of the effects of TMAO on aging and neurodegeneration, related to dietary intake and gut health
Lee et al. (2022)
Study of gut-liver-cerebral interactions in Parkinson’s disease
Trapecar et al. (2021)
Blood-Brain Barrier
Modeling of the blood-brain barrier using assembloids with brain and blood vessel organoids
Dao et al. (2024)
4.2. Organs-on-chip: simulating the dynamic gut-brain environment
Organ-on-Chip (OoC) models, also known as Microfluidic platforms, integrate lab-on-chip technology with 3D organotypic cultures to replicate the pathophysiological complexity of the microbiota-gut-brain axis. OoC models utilize various culture channel surfaces and require less media, offering flexibility in both single-organ and more intricate multi-organ configurations (Aldea et al., 2019). These platforms feature thin culture chambers that allow for continuous perfusion of culture media, making them ideal for imaging and quantitative assays, as they permit the harvest of a sufficient number of cells. Additionally, OoC devices can replicate key functional aspects of tissue physiopathology, enabling the evaluation of food bioactive compounds or therapeutic agents and their effects on targeted tissues. They offer the advantage of reducing research costs and increasing throughput compared to animal models, thereby addressing ethical concerns. OoC systems have the potential to accurately model the microbiota-gut-brain microenvironment, closely mimicking the physiological features seen in vivo (Aldea et al., 2019). These platforms recreate in vivo organ physiology and function in a controlled in vitro setting. In addition, OoCs provide exceptional control and independent spatiotemporal tuning of key factors within the cell system (e.g., pH, oxygen), alongside automated in situ monitoring and sample analysis, as well as downstream analysis. They also offer the potential to study cell-cell and cell-niche interactions with high precision.
The advantages of physiologically relevant, animal-free models have recently been harnessed for the development of gut-on-chip systems. These systems have demonstrated the successful replication of region-specific structures and functions of both the small and large intestines using organ-on-chip technology (Özkan et al., 2024). These approaches offer significant improvements over traditional in vitro models by allowing for the continuous supply of nutrients and removal of waste, incorporation of vascular and immune system components, as well as intestinal flora (Bein et al., 2018; Ronaldson-Bouchard & Vunjak-Novakovic, 2018). Additionally, some models apply mechanical deformations to simulate gut peristalsis (Jing et al., 2020). Most recent OoC platforms are capable of maintaining long-term co-culture of live human intestinal epithelial cells alongside a complex microbial community (Dickson, 2019).
Other strategies focus on integrating sensing units for real-time monitoring of critical parameters, such as oxygen levels (Jalili-Firoozinezhad et al., 2019) and barrier integrity (van der Helm et al., 2019) which are essential for studies on the gut-microbiota-brain axis.
Min et al. (2022) introduced a pathomimetic “Leaky Gut-Chip,” which successfully models increased epithelial permeability and intestinal inflammation to evaluate the therapeutic potential of probiotics. This advanced gut-on-chip platform replicates essential features of the intestinal microenvironment, including biomechanical forces and oxygen gradients. By introducing pro-inflammatory cytokines (TNF-α and IL-1β), the model reliably induced prolonged epithelial barrier dysfunction. Probiotic treatments, such as Lactobacillus rhamnosus GG and the multi-species mixture VSL#3, significantly improved barrier function, restored tight junction protein localization, and reduced inflammation. The “Leaky Gut Chip” provides a robust tool for investigating intestinal barrier dynamics and evaluating the efficacy of probiotic interventions, with broader implications for gut-brain axis research and the potential impact of probiotics on systemic health (Min et al., 2022).
Sedrani et al. (2023) developed the human-microbial crosstalk gut-on-chip model by incorporating iPSCs-derived enteric neurons into the gut compartment. This enhanced model enables the co-culture of bacterial, epithelial, and neuronal cells within microfluidic channels that are separated by semi-permeable membranes. While the different cell types remain physically separated, they can still communicate via soluble factors, allowing for independent study of each cell type. This setup can offer novel insights into how the gut microbiome influences enteric neuronal cells, providing a crucial foundation for investigating the gut microbiome-nervous system axis (Sedrani et al., 2023).
Similarly, OoC technology has been employed to create more advanced models of the blood-brain barrier (BBB) and/or the brain (Maoz et al., 2018; Vatine et al., 2019) demonstrating significant potential for evaluating whether drug candidates, including bioactive compounds from food or specific metabolites, can effectively cross the BBB and reach their target sites in the brain.
Park et al. (2019) developed a blood-brain barrier (BBB) model on a chip, incorporating induced pluripotent stem cell (iPSC)-derived human brain endothelial cells differentiated under hypoxic conditions, along with primary human pericytes and astrocytes. This model successfully maintained in vivo-like BBB barrier integrity and transport functions for up to one week (Park et al., 2019).
Mir et al. (2022) developed a BBB-OoC platform designed to replicate the functionality of the in vivo BBB for the study of neurodegenerative diseases and drug permeability testing (Mir et al., 2022). This platform has demonstrated significant value in improving the understanding of neurodegenerative disease mechanisms and in facilitating drug testing. A notable feature of this model is the integration of biosensors and detection systems, which allows for real-time biomarker monitoring and automated drug permeability analysis, enhancing the platform’s efficiency and utility. In the context of gut-brain axis and nutrition research, this platform is particularly relevant for investigating how dietary components and gut-derived metabolites influence brain health via the BBB. The gut microbiota plays a crucial role in metabolizing nutrients into bioactive compounds which can impact both BBB integrity and brain function. The integration of biosensor technology into the BBB-OoC platform provides a valuable tool for studying the effects of specific nutrients or dietary interventions on BBB permeability, opening new avenues for exploring the relationship between diet, gut health, and neuroinflammation.
There is significant interest in functionally linking individual OoCs through their endothelial or vascular channels in an in vivo-like sequence, creating multi-organ OoC systems that replicate vascular perfusion and inter-tissue communication. This approach has recently proven particularly useful for studying microbiota effects on the brain. In one study, OoCs mimicking the gut-liver-kidney-brain axis were coupled to investigate the toxicity of microbiome-derived metabolites, such as trimethylamine (TMA) and Trimethylamine N-oxide (TMAO). TMA is initially produced by gut microbiota during the metabolism of dietary compounds such as choline, l-carnitine, and betaine, which are commonly found in foods like red meat, eggs, and dairy. After TMA is absorbed into the bloodstream, it is transported to the liver, where it is oxidized into TMAO, which is associated with increased risk for major adverse cardiovascular events and death in humans. The study demonstrated that TMAO can cross the blood-brain barrier (BBB) and reach the neurovascular unit (Vernetti et al., 2017).
Trapecar et al. (2021) developed a modular GBA chip to simulate interactions between the gut, liver, and brain. This platform incorporates a central circulatory system that connects each module along the gut-liver-brain axis. By utilizing this system, they successfully replicated in vivo-like behaviour within the brain module and demonstrated that microbiome-associated SCAFs enhanced the expression of pathways related to pathology in patients with Parkinson’s disease (Trapecar et al., 2021).
Kim et al. (2024) recently investigated the role of microbe-derived metabolites and exosomes on neurodevelopment and neurodegenerative disorders, using neurons derived from human iPSCs within a GBA chip. Their research revealed that these microbial products significantly influenced neuronal growth, maturation, and synaptic plasticity (N. Y. Kim et al., 2024). In the context of nutrition these findings suggest that advanced models, such as GBA chips, can play a crucial role in exploring how dietary interventions modulate the gut microbiota. By using these models, researchers can assess how specific compounds derived from diet or supplements promote the production of beneficial microbial metabolites and exosomes. These microbial products, in turn, could influence brain health, providing a basis for potential therapeutic strategies aimed at neurodevelopmental and neurodegenerative disorders. Fig. 2 shows a typical example of a multi-organ-on-a-chip system for the gut-brain axis within the context of nutrition research.
Download: Download high-res image (1MB)
Download: Download full-size image
Fig. 2. A representative example of a multi-organ-on-a-chip system for studying the gut-brain axis. (A) Illustration of the anatomical components of the gut-brain axis (GBA), incorporating the gut microbiome. Communication between the brain and gut occurs bi-directionally via the vagus nerve, immune pathways, and the neuroendocrine system (B) Illustration of the micro-anatomy of the GBA: (bottom) microbiota within the intestinal lumen and mucosal layer interact with epithelial cells (e.g., enterocytes). of the intestinal brush border, vascularized by capillaries that transport red blood cells and lymphocytes via systemic circulation to the blood-brain barrier, which interfaces with neurons, pericytes, astrocytes, microglia, and oligodendrocytes in the brain. (C) Schematic of a Multi-Organ Chip, linking multiple MPSs, including (bottom) microbiota-on-a-chip, gut-on-a-chip (enterocytes), immune-on-a-chip (macrophages and lymphocytes), blood-brain barrier-on-a-chip (cerebrovascular endothelial cells), and (top) brain-on-a-chip (neurons, microglia, and astrocytes embedded in hydrogel). Reproduced from (Boylin et al., 2024): CC BY 4.0. (For interpretation of the references to colour in this figure legend, the reader is referred to the Web version of this article.)
OoC systems hold significant promise for advancing research on the GBA and nutrition by enhancing our understanding of complex interactions between these systems and their role in health and disease. These platforms can facilitate the development of novel therapeutic strategies and nutritional interventions. However, despite their potential, several limitations remain to be addressed. Current microfluidic OoC models, while capable of simulating various human phenotypes and organ-specific responses, do not yet fully replicate the structural and functional complexities of human tissues. For example, in gut-on-chip models, the four-layered structure of the intestinal wall is not fully reproduced, even though these layers are essential for accurately modeling conditions where the gut-brain axis is involved, such as in inflammatory and metabolic diseases. To better study such conditions, it is crucial for gut-on-chip systems to integrate all four layers of the intestinal wall, capturing their unique characteristics and interactions. This enhancement will be pivotal in creating more advanced models for gut-brain axis and nutrition research. Moreover, the development and maintenance of OoC models remain technically challenging and resource-intensive. These systems demand a multidisciplinary approach, requiring expertise in microfabrication, bioengineering, and biology, which increases their complexity. Additionally, the initial costs for establishing these platforms are often high, further complicating their widespread adoption (Candarlioglu et al., 2022). Addressing these challenges is crucial to fully harness the potential of OoC models for advancing gut-brain axis research in the field of nutritional science.
While organs-on-chip technology enables the modeling of the gut-brain axis with improved physiological relevance, further insights into molecular mechanisms require advanced analytical approaches. Omics sciences, including genomics, transcriptomics, and metabolomics, allow for large-scale data acquisition to elucidate the molecular and microbial influences on gut-brain interactions.
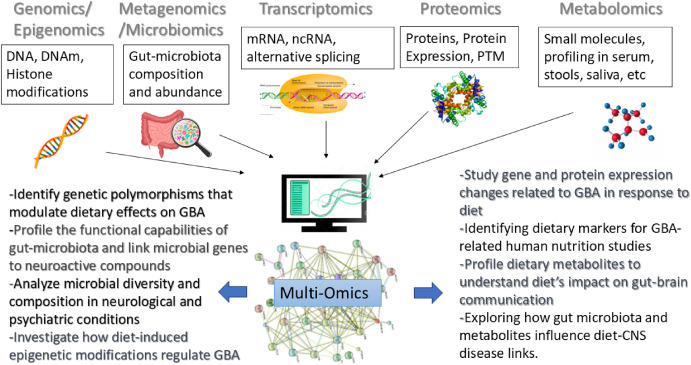
4.3. Omic sciences: comprehensive analysis of gut-brain interactions
Modern nutritional research is beginning to harness the potential of omics sciences – including genomics, transcriptomics, proteomics, metabolomics, metagenomics, microbiomics, and epigenomics – to better understand the molecular and systemic complexity of the GBA. This emerging capability has been significantly enhanced by advancements in next-generation sequencing (NGS), mass spectrometry, nuclear magnetic resonance, high-throughput platforms, and bioinformatic tools. The evolution of these technologies is paving the way for comprehensive and simultaneous analysis of large-scale datasets with unprecedented precision and depth, with great potential to offer new insights into the intricate connections between diet, gut health, and brain function.
4.3.1. Genomics in gut-brain axis studies
Genomics encompasses the study of the complete DNA sequence of an organism, cell, or tissue, offering insights into genetic predispositions that influence how nutrients affect the gut microbiota and neurophysiological functions. Advances in next-generation sequencing (NGS) enable the simultaneous sequencing of numerous short nucleic acid fragments, delivering rapid, high-throughput data at reduced costs (Scarano et al., 2024). When integrated with genome-wide association studies (GWAS), NGS has been instrumental in identifying novel genomic variants, such as genetic polymorphisms, enhancing our understanding of the interaction between dietary exposures and genetic variation in humans. Genetic polymorphisms can modulate the impact of dietary components on the GBA. GWAS have helped identify genetic variants linked to microbiota composition, highlighting potential genetic factors that shape host-microbiome interactions (Blekhman et al., 2015).
Weiming et al. (2023) conducted a genome-wide pleiotropic association study to explore the shared genetic basis between gastrointestinal diseases and psychiatric disorders, identifying numerous shared genetic variants, pleiotropic loci, and candidate genes enriched in traits and tissues relevant to the gut-brain axis. Their findings highlight pathways related to cell adhesion, synaptic function, and immune differentiation, supporting a common genetic foundation underlying the gut-brain axis and suggesting potential targets for treating both GI and psychiatric conditions (Gong et al., 2023).
Zhou et al. (2023) used GWAS data to examine the potential influence of gut microbiota on brain structure, specifically looking at cortical surface area and thickness across various brain regions. They found that higher predicted levels of certain gut microbial taxa, such as Mollicutes and Tenericutes, were associated with increased surface area in the orbitofrontal cortex – a region implicated in neuropsychiatric disorders (M. Zhou et al., 2023).
Genomic approaches can help personalize nutritional interventions to optimize GBA function. Understanding these genetic components allows for more targeted dietary recommendations, enhancing individual outcomes for both gut and brain health.
4.3.2. Metagenomics: profiling the gut microbiome
Metagenomics allows researchers to analyze the collective genetic material of microbial communities residing in the gut. This field has expanded our understanding of the gut microbiome’s contribution to GBA function. Through metagenomic sequencing, the functional capabilities of gut microbiota can be inferred, linking specific microbial genes to neuroactive compound production (Tilocca et al., 2020). Metagenomics could be instrumental in identifying microbial pathways affected by diet, helping to design specific interventions to enhance gut microbiome profiles that support optimal brain function.
4.3.3. Microbiomics: gut microbial composition and GBA
Microbiomics is a key omics approach for analyzing the diversity and abundance of microbial species in the gut. This technology focuses on profiling the structure of microbial communities to understand their role in the GBA.
In ongoing clinical investigations into the microbiomes of neurological and psychiatric patients, a substantial volume of taxonomic data – primarily centered on bacterial species identified in fecal samples – is continuously being generated. Using these phylogeny-based biomarkers, researchers can identify disease-associated microbes, including both commensal and pathogenic species. This data facilitates the assessment of microbial community abundance and composition and enables predictive analyses of microbial functions based on the expression of specific genes, RNA, and proteins, as well as their involvement in molecular and cellular pathways. By comparing microbial diversity and the presence of unique taxa between individuals with specific disorders and matched control groups, alterations in disease-associated microbiomes have been identified across a range of psychiatric and neurological conditions, including Parkinson’s Disease, multiple sclerosis, Autism Spectrum Disorder, schizophrenia, Huntington’s Disease, Attention-Deficit Hyperactivity Disorder, Major Depressive Disorder, and Alzheimer’s Disease (Severance, 2024). Microbiomic studies may help elucidate how specific dietary interventions promote the growth of beneficial microbes that positively influence mood and cognitive function (J. Zhou et al., 2024).
4.3.4. Transcriptomics and proteomics: gut and brain response to nutrition
Transcriptomics, enabled by advanced technologies such as Single-Cell RNA Sequencing (scRNA-seq), focuses on analyzing RNA expression to provide detailed insights into how dietary factors influence gene activity within both gut and brain tissues. Ghosh et al. highlighted how a Mediterranean diet can influence gut microbiota and its related gene expression changes in the context of human subjects, linking these changes to improved inflammatory markers and potential cognitive benefits (Ghosh et al., 2020).
Proteomics, the large-scale study of proteins, enables the assessment of protein expression changes in response to dietary alterations. This technology could provide critical information on how diet impacts both gut barrier proteins (such as tight junctions) and neuroactive peptides (Ganesh & Hettiarachchy, 2012).
4.3.5. Epigenomics: nutritional modulation of gene expression
Epigenomics investigates how external factors, including diet, can lead to modifications in gene expression without altering the underlying DNA sequence. Such modifications, including DNA methylation and histone acetylation, can have profound impacts on GBA function. Dietary components like polyphenols, vitamins, and short-chain fatty acids have been shown to induce epigenetic changes that regulate inflammation and neurotransmitter synthesis (Berni Canani et al., 2012).
Understanding these modifications helps researchers develop nutritional interventions that can potentially mitigate the risk of neuropsychiatric disorders through the modulation of epigenetic marks. Epigenomic research is especially relevant for developing preventive strategies in at-risk populations, as it provides a mechanism to understand how early-life nutrition can influence long-term brain health and behaviour (Louwies et al., 2019).
4.3.6. Metabolomics: linking diet, microbial metabolites, and mental health
Metabolomics, which involves the profiling of small-molecule metabolites, provides direct insights into the biochemical consequences of diet on the GBA. Traditionally, Nuclear Magnetic Resonance (NMR), Proton Nuclear Magnetic Resonance (1H NMR) Spectroscopy, and Mass Spectrometry (MS) have been the primary technologies applied in metabolomics to explore the biochemical pathways influenced by gut microbiota. The primary analytical platforms for metabolomic analysis of the microbiota-gut-brain axis include liquid chromatography coupled with mass spectrometry (LC-MS), which enables quantification of various organic acids, including short-chain fatty acids (SCFAs), bile acids, and their derivatives. Gas chromatography coupled with mass spectrometry (GC-MS) also serves as a robust technique for metabolite profiling. Common samples and matrices for metabolomic analyses encompass plasma, serum, cerebrospinal fluid, stool, urine, intestinal biopsy specimens, and brain tissue (Konjevod et al., 2021).
Recent advancements in these technologies, particularly Matrix-Assisted Laser Desorption Ionization Time Of Flight (MALDI-TOF), Secondary Ion Mass Spectrometry (SIMS), and Fourier transform ion cyclotron resonance MS, along with emerging methods for metabolite identification, have facilitated the quantitative, non-invasive analysis of metabolites in body fluids and tissues. These advancements are crucial for identifying metabolic changes influenced by diet and microbial activity, which may affect brain function and contribute to neurodevelopmental and neurodegenerative processes through the gut-brain axis (Tilocca et al., 2020).
Diet-derived metabolites such as SCFAs, bile acids, and tryptophan metabolites (including serotonin) are key mediators in gut-brain communication (Berni Canani et al., 2012). Metabolomics can assess how different diets – such as ketogenic, Mediterranean, or high-fat diets -affect metabolite profiles and, in turn, influence brain health.
One prominent example is the role of SCFAs, such as butyrate, which is produced by microbial fermentation of dietary fibers. Butyrate has anti-inflammatory properties and modulates the gut-brain barrier, potentially reducing neuroinflammation associated with mood disorders (Dalile et al., 2019). Metabolomics also aids in understanding the mechanisms by which specific nutrients or food patterns contribute to changes in metabolite levels, offering targets for nutritional interventions that support both gut and brain health.
4.3.7. Multi-omics integration: a holistic approach to the gut-brain axis
The integration of multiple omics approaches, known as multi-omics, provides a comprehensive understanding of the GBA. By combining genomics, metagenomics, transcriptomics, proteomics, microbiomics, epigenomics, and metabolomics, researchers can gain a holistic view of how diet affects the gut and brain simultaneously. For example, a recent multi-omics study by Li, Liu, et al. (2023) investigated the role of altered gut microbiota and microbiota-derived metabolites in modulating the association between unbalanced dietary intake and gastrointestinal symptoms in children with autism spectrum disorder (Li, Liu, et al., 2023).
A study combining microbiomics and metabolomics investigated the role of gut microbial composition and metabolites in schizophrenia. Using 16 S rRNA gene sequencing and untargeted LC-MS, the study profiled gut bacteria and microbial metabolites in healthy controls, acute patients, and remission patients. The results revealed significant microbial disturbances and abnormal metabolites in patients, with certain bacterial taxa and metabolites correlating with schizophrenia severity. These findings suggest that gut microbiota and their metabolites may interactively influence schizophrenia’s pathophysiology, providing insights into the brain-gut axis in central nervous system disorders (Shi et al., 2023). Fig. 3 summarizes the primary applications of omics sciences in analyzing GBA interactions.
Download: Download high-res image (910KB)
Download: Download full-size image
Fig. 3. Overview of Omics Approaches and their Applications in GBA studies within nutritional research. Each omics discipline provides insights into specific aspects of GBA interactions, ranging from genetic influences and microbial composition to metabolite profiling and protein expression. Integrating multiple omics approaches (multi-omics) allows for a comprehensive understanding of how dietary components impact the gut-brain axis in both health and disease.
Despite the significant opportunities that omics and multi-omics approaches offer for advancing gut-brain axis research in the context of nutrition, several challenges must still be addressed to fully harness the potential of integrating high-throughput data across different molecular layers. These challenges include the heterogeneity among omics technologies, managing missing data, the complexity of interpreting multilayered systems models, and issues related to data annotation, storage, and computational resources (Tarazona et al., 2021). Analysing and interpreting these large-scale datasets necessitate advanced computational models. In silico approaches, including AI-driven predictions and machine learning, provide powerful tools for simulating gut-brain interactions and optimizing experimental models.
4.4. Computational models
Computational models, also known as in silico models, integrate mathematical and computational methods, including quantitative structure–activity relationships (QSAR), artificial intelligence (AI), machine learning (ML), and others. They have become increasingly valuable tools for studying the GBA (Hawkins et al., 2020; Mohammad et al., 2022; Shokri Garjan et al., 2023). These models could enable researchers to simulate complex biological interactions between the gut microbiome, dietary factors, metabolism, and brain function, thereby offering insights that complement experimental studies.
Khan Mohammad et al. (2023) created a computational model to investigate the GBA in autism spectrum disorder (ASD), utilizing constraint-based metabolic modeling (CBM)alongside whole-body pharmacokinetics (PBPK) modeling to explore metabolic irregularities in both gut and brain regions. This model identified essential metabolic pathways – such as carbon and vitamin metabolism within the gut and mitochondrial energy and amino acid pathways in the brain – that are linked to ASD. By simulating dietary and probiotic strategies, the study showed that a high-fiber diet and the incorporation of specific probiotics could support microbiota balance and reduce oxidative stress within the gut-brain axis. This comprehensive framework illustrates the potential of integrating CBM and PBPK modeling to pinpoint ASD therapeutic options and offers a promising avenue for GBA research in nutrition-based interventions (Mohammad et al., 2022).
Kovtun et al. (2020) examines the presence and distribution of genes responsible for synthesizing enzymes that produce neuroactive compounds in 147 gut metagenomes from healthy individuals (sourced from the Human Microbiome Project) and a synthetic metagenome assembled from 508 bacterial genomes. Using a catalog of orthologs for 17 key enzymes and a specialized search algorithm, the analysis identifies seven bacterial genera with the highest abundance of enzyme genes and the eight most frequently observed enzymes. These findings suggest that the identified “core” genera and enzymes represent a metagenomic signature of the neurometabolic potential in healthy human gut microbiota, demonstrating the potential of computational in silico approaches in elucidating gut-brain axis mechanisms (Kovtun et al., 2018).
Quinn-Bohmann et al. (2023) used a microbial community-scale metabolic modeling approach to predict individual-specific profiles of SCFA production in the gut, aiming to understand how dietary, prebiotic, and probiotic inputs impact SCFA levels. The accuracy of these predictions was validated using in vitro, ex vivo, and human cohort data, showing significant associations between SCFA production predictions and clinical markers of cardiometabolic and immune health (Quinn-Bohmann et al., 2024). Given the influence of SCFAs on the gut-brain axis (GBA) via immune modulation, inflammation control, and neurotransmitter synthesis, this model holds potential for GBA studies by identifying dietary interventions that may enhance SCFA-mediated brain health benefits.
Among computational methods with potential applications for studying the gut-brain axis in nutrition research is the digital twin (DT). A DT is a virtual model designed to replicate the characteristics and behavior of physical entities, both living and nonliving, enabling real-time simulation and analysis. Acting as a dynamic representation, a DT mirrors human cells, tissues, organs or microenvironments, adapting continuously to real-time data inputs and forecasting future scenarios based on those changes (Vallée, 2023).
To advance the discovery, understanding, and management of GBA disorders, the OnePlanet research center is creating digital twins specifically focused on the GBA. These digital twins integrate innovative sensors with AI algorithms to deliver descriptive, diagnostic, predictive, or prescriptive feedback (Meystre et al., 2023).
Computational models play a critical role in in vitro to in vivo extrapolation (IVIVE) by integrating data from controlled laboratory experiments and translating them into predictions for more complex biological systems (Fragki et al., 2023). In the context of GBA and nutrition research, these models help bridge the gap between in vitro findings and in vivo responses. For example, in vitro gut-on-chip systems enable detailed mechanistic exploration of microbial metabolism, nutrient interactions, and food contaminant toxicity. Computational simulations can then leverage this data to predict systemic effects in humans, such as the impact of dietary interventions and microbiome shifts on both local gut environments and brain function. This integrative approach offers a comprehensive framework to study the interactions among diet, food-derived contaminants, gut health, and brain activity, providing a more accurate representation of in vivo physiological processes.
The primary limitation of computational models is their reliance on the data they are trained on or tasked to analyze; their effectiveness is directly tied to data quality, and performance declines without regular updates. Therefore, continuously supplying and refreshing these models with new, human-relevant data is essential for maintaining accuracy and relevance.
5. Conclusion and future perspectives
This review emphasizes the transformative potential of NAMs in enhancing research on the GBA within the field of nutritional science. In recent years, there have been significant advancements in scientific research aimed at developing human-centered methodologies that reduce the dependence on animal models. Regulatory changes, such as the FDA Modernization Acts 2.0 and 3.0, have removed the requirement for animal testing in drug development and established a structured framework to support non-animal methodologies (J. J. Han, 2023). Likewise, the EFSA 2027 Strategy encourages a shift towards NAM-based food risk assessments, reinforcing a commitment to ethical and human-relevant models (Cattaneo et al., 2023). On a global scale, initiatives like the creation of a comprehensive international NAMs database aim to provide standardized references for selecting suitable methodologies (Storz & Dean, 2024). In light of these developments, NAMs are anticipated to gain traction across various research sectors. Their use in GBA studies within nutritional science is particularly significant, as the interactions between the gut microbiome, diet, and brain are complex and necessitate advanced models that can accurately reflect human-specific mechanisms. NAMs present an opportunity to improve human relevance and translatability while simultaneously decreasing reliance on traditional animal models. In Europe alone, around 45,000 animals are utilized each year for regulatory testing in the food sector (de Boer et al., 2020). Increasing public and regulatory scrutiny regarding animal testing further highlights the need to tackle both the scientific and ethical challenges present in biomedical research. The Directive, 2010/63/EU, which aims for the eventual replacement of all animal research with non-animal alternatives, underscores this urgent necessity (Directive, 2010, Recital 10).
5.1. A balanced comparison: NAMs vs. animal models
While NAMs provide significant benefits, such as greater human relevance, improved predictive accuracy, and ethical sustainability, traditional animal models are still commonly used because of their accessibility, regulatory acceptance, and established infrastructure. Many universities, hospitals, and research institutions do not have the technical or financial means to implement NAMs on a large scale, which restricts their immediate application (Fig. 4). Therefore, even with the increasing interest in alternative methods, traditional models are likely to remain important, especially for validating NAM-generated data against human studies and fostering trust within the scientific community. One major drawback of NAMs is that no single model can fully replicate human physiology, just as no animal model can. However, a tiered or integrated approach that merges NAMs with human observational and intervention studies could improve both translational accuracy and scientific reliability in GBA and nutrition research (Andersen et al., 2019).
Download: Download high-res image (536KB)
Download: Download full-size image
Fig. 4. Comparison of Traditional Animal Models and NAMs in GBA and nutrition research. Traditional animal models benefit from regulatory acceptance, standardized protocols, and broad availability but are limited by low human relevance, ethical concerns, and resource-intensive requirements. In contrast, NAMs offer higher human relevance, improved predictive accuracy, and ethical sustainability but face challenges related to regulatory acceptance, validation, and standardization. The symbolic scale illustrates the trade-offs between these approaches, emphasizing the need for standardization, validation, and addressing biases favoring traditional methods to facilitate the broader adoption of NAMs in regulatory and scientific frameworks.Challenges to Adoption and Feasibility Considerations.
Despite their acknowledged benefits, various challenges still impede the broader use of NAMs in biomedical research. These obstacles arise from scientific, technical, legislative, and economic factors, along with a cultural preference for traditional methods. For example, as of 2022, fewer than 10 % of top nutrition and dietetics journals explicitly endorsed human-based, non-animal research (Storz & Dean, 2024). Moreover, deep-seated biases exist within the scientific community, where studies advocating for NAM-based methods often encounter reviewer requests for animal validation, which restricts their acceptance in both publications and funding avenues (Ingber, 2020). To overcome these challenges, focused education and awareness efforts are needed, engaging researchers, students, journal editors, funding bodies, and policymakers. Training programs and outreach initiatives can highlight the scientific benefits of NAMs while also increasing familiarity with these methods in academic and regulatory environments. Additionally, investing in technical infrastructure is vital, as many research institutions still lack the necessary facilities to effectively implement NAMs.
While NAMs have shown better performance in various applications (Barrile et al., 2018; Hartung, 2019; Nieskens et al., 2021), their validation process remains resource-heavy. For instance, a qualification study involving 870 liver-on-chips for drug-induced liver injury (DILI) achieved 87 % sensitivity, successfully identifying hepatotoxic drugs that traditional animal models missed, which were later associated with 242 DILI-related fatalities (Ewart et al., 2022). Such studies are essential for establishing scientific credibility and gaining regulatory acceptance for NAMs.
5.2. A feasible and sustainable transition
Investment in NAMs often faces limitations due to perceived high risks, even though they hold the promise of enhanced scientific accuracy, financial efficiency, and greater relevance to human health. To promote their adoption, it is crucial to establish a clear regulatory framework that connects risk capital to viable NAM-based products. Furthermore, policies that align with societal expectations—such as fostering economic growth, encouraging innovation, and minimizing animal testing—will be instrumental in facilitating this shift (Parvatam et al., 2024). To fully realize the potential of NAMs in GBA and nutrition research, it is essential to enhance their feasibility. This requires collaboration across sectors, involving researchers, policymakers, industry stakeholders, and regulatory bodies, to address scientific, technological, and regulatory challenges. Increased investment in research and development, along with targeted funding for validation and standardization, will be vital for making NAMs practically applicable. By promoting interdisciplinary cooperation and ensuring ongoing regulatory and financial backing, NAMs can be seamlessly integrated into mainstream research (Parvatam et al., 2024). While a complete replacement of traditional models may not be feasible in the near term, NAMs present a scientifically sound, human-relevant, and ethically sustainable alternative that has the potential to transform GBA and nutrition research.
Abstract Background The gut-brain axis is a complex communication network that connects the gastrointestinal system with the central nervous system, significantly influencingRead More
New research has estimated that, given the right support, innovative ways of producing everyday foods using fermentation could add £9.8 billion (€11.2Read More
Get in touch Contact
Have questions about our precision nutrition platform, research collaboration, or global market access? Our team is here to support physicians, partners, and innovators